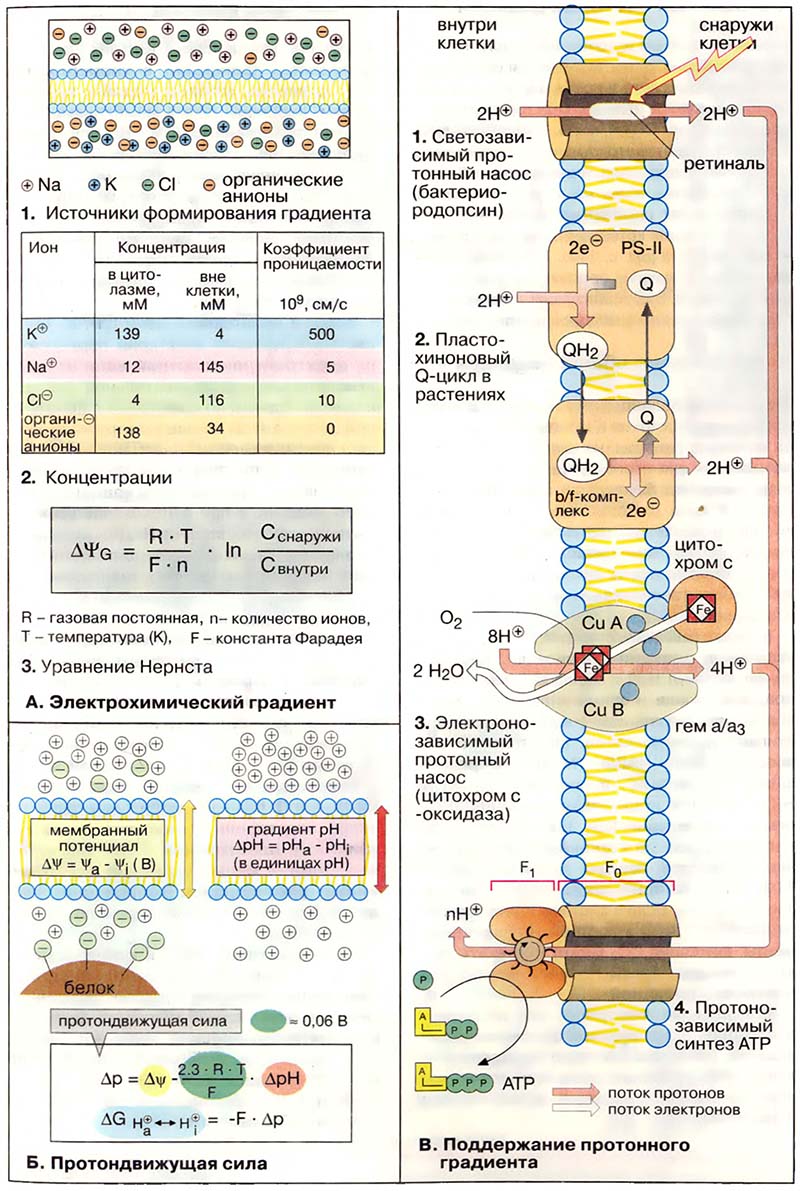
В то время как искусственная липидная мембрана для ионов практически не проницаема, биологические мембраны содержат «
ионные каналы», по которым отдельные ионы избирательно проникают через мембрану. Проницаемость и полярность мембраны зависят от
электрохимического градиента, то есть от концентраций ионов по обе стороны мембраны (
концентрационного градиента) и от
разности электрических потенциалов между внутренней и внешней сторонами мембраны (
мембранного потенциала).
В состоянии покоя клеток мембранный потенциал (
потенциал покоя) составляет от −0,05 до −0,09 В, то есть на внутренней стороне плазматической мембраны преобладает избыток отрицательных зарядов.
Потенциал покоя обеспечивается прежде всего катионами Na
+ и K
+, а также органическими анионами и ионом Cl
- (1). Концентрации снаружи и внутри клетки и коэффициенты проницаемости этих ионов приведены в таблице (2).
Распределение ионов между внешней средой и внутренним объёмом клетки описывается
уравнением Нернста (3), где ΔΨ
G — трансмембранный потенциал (в вольтах, В), то есть разность электрических потенциалов между двумя сторонами мембраны при отсутствии транспорта ионов через мембрану (
потенциал равновесия). Для одновалентных ионов при 25°С множитель RT/Fn равен 0,026 В. Вместе с тем из таблицы (2) следует, что для ионов K
+ ΔΨ
G примерно равно −0,09 В, т. е. величина того же порядка, что и потенциал покоя. Для ионов Na
+, напротив, ΔΨ
G ≈ +0,07 В, то есть выше, чем потенциал покоя. Поэтому ионы Na
+ поступают в клетку при открытии Na
+-канала. Неравенство концентраций ионов Na
+ и K
+ постоянно поддерживается
Na+/K+-АТФ-азой при расходовании АТФ.
®